縧蟲感染症
1. 疾病描述
病原:縧蟲(Cestoda)
病原環境:海水、半淡鹼水及海水(marine、brackish & freshwater)
學名:例︰
(1)廣節裂頭縧蟲(
Diphyllobothrium latum)
(2)腸舌狀縧蟲(
Ligula intestinalis)
(3)鱊頭槽縧蟲(
Bothriocephalus acheilognathi)
(4)九江頭槽縧蟲(B.
gowkongensis),同義於(3)之命名
(5)其他縧蟲(Other cestodes)
病名(及俗名):(1)鱊頭槽縧蟲稱為亞洲魚縧蟲(Asian fish tapeworm)或錦鯉縧蟲(Koi tapeworm)
(2)腸舌狀縧蟲稱為舌狀縧蟲病(Ligulosis)
最早發現者:例︰
(1)腸舌狀縧蟲 ( L.
intestinalis );Linnaeus 1758, Gmelin, 1790.
(2)鱊頭槽縧蟲 ( B.
acheilog nathi ), Yamaguti, 1934.
(3)廣節裂頭縧蟲 ( D.
latum ), Linnaeus, 1758.
OIE狀況:未表列(Not Listed)
病原摘要:
以整個縧蟲體部之組成要素來做機要性敍述,其後在疾病特性中,再將個別縧蟲體舉例陳述。
一般而言扁平動物內縧蟲綱無脊椎動物,約有3,000種,體長約1公分至15公尺,世界性分佈;寄生在某些無脊椎動物及各類脊椎動物如人,魚及家畜等動物之肝及消化道內(腸為主),引起縧蟲病(cestodiasis);在蟲體生活史中,有的只有一個宿主,有的除終宿主外,還需有1至2個中間宿主。
縧蟲為雌雄共同體之內寄生蟲,長而扁平。無體腔及消化道,蟲體包含有︰(1)頭節(scolex或head),通常具有吸盤(acetabulum或sucker)、鉤(hook),或吸溝(bothrium),見圖1。(2)蟲體(strobilum),由極多之節片(proglottikes或segments)構成。(3)頸(neck),介於頭節及體節間之無分節部份。每一節片通常含有1或2對之雌雄生殖器。蟲體體長自數公分至數十公尺不等,頭節一般為球形,具2至4個肌肉質吸盤,有的其上具鉤。頭之前端有一突起,稱吻(rostellum),其上具1至數排之鉤。有時鉤甚小而不易發現。蟲體由節片構成,自頭部往後生出。前側年青且體小,往後則逐漸成熟寬大。節片之數目隨縧蟲種類而異。擬葉目縧蟲之頭節不具鉤,代之以兩片葉狀之吸溝(bothrium)。縧蟲之組織似吸蟲,體表有角質層,特稱覆皮(tegument)。覆皮異於一般角皮,含多種消化酵素,具吸收作用。電子顯微鏡顯示覆皮表層被覆纖毛(cilia),與一般腸細胞表層纖毛同。蟲體靠前端之節片最先出現雄性生殖系,再往後側才出現雌性生殖系。凡是生殖系已達成熟,且開始進行生殖作用之節片均稱為成熟節片(mature segment)。當蟲卵受精後(節片間相互受精或單獨節片自體受精)生殖器便退化,只留下充滿受精卵之子宮。此節片特稱妊娠節片(gravid segment),其於成熟後節片隨寄主糞便排出體外。擬葉目縧蟲之妊娠節片則於雌生殖孔下方另具排卵孔,直接將蟲卵排出體外,當卵排完後後衰老之節片再分串排出體外。
縧蟲體主要由成鏈之節片(proglottids)組成,每一節片內含有一組或多組生殖器官,而節片由無性生殖之出芽長出(asexual budding),全部節片組成縧蟲體(strobila),這些節片性稱為polyzoic;某些族體,特別是魚類縧蟲,有時僅存在一片體(segment),稱為monozoic;而頭節(scolex)是縧蟲生活史和生長的重要部份,並常為分類依據;見圖1。位於體前端,上有各種附著器官,包括吸盤(suckers)、鈎(hooks)、曲溝(grooves),棘(spines)、腺體區(glandular area)。有些種類頭節退化或發育不全,有些種類之頭節還有腺體存在,頭節內有神經及感覺末稍,排泄管亦集中聯結頭節內;依Jones A, Bray RA及Khalid F等人於1994年之整理分類,在縧蟲綱(Cestada)下,分有14目縧蟲,在魚類普遍重要的有假葉目(Pseudophyllidea Carus, 1863)、圓葉目(Cyclophyllidea)及鯉蠢目(Caryophyllidea Bea, in Carus, 1863),假葉目及圓葉目之俗名分別為擬縧蟲(Pseudotapewoum)及真縧蟲(true tapeworm);前者屬水生食物環(aquati food chains),而後者屬陸生食物環(terreserial food chains),假葉目下縧蟲之種類甚多,多寄生於魚類,只有目下裂頭科(Diphyllo- bothriidal)寄生哺乳動物。所有魚類縧蟲均是雌雄同體,在蟲體的生活史中至少有一個中間宿主,大多數為非脊椎動物如寡毛類(oligochceetes, 顫蚓)、橈腳類(copepoda, 劍水蚤)、端腳類(amphipoda)。一般分類依據大致如下︰詳細情形可參閱,魚類寄生蟲與寄生蟲病,此書由張劍英等編著。
1(2)縧蟲體無內部分節(internal segmentation),有一組生殖器官………………………………………………………鯉蠢目(Caryophyllidea)
2(1)縧蟲體具有內部分節,超過一組以上之生殖器官
3(4)頭節不具真吸盤(true suckers)、吸溝(bothria)、裂片(bothridia)、觸鬚(tentacles)、不具外分節(external segmentation)……窄溝目(Spathebtriidea)
4(3)頭節上含有固著器型(holdfast type)(3),外分節經常遠距分離
5(6)頭節含有二節吸溝……………………………………假葉目(Pseudphyllidea)
6(5)缺乏吸溝,頭節排列成四個圓狀吸盤(armed with four rounded suckers)………………………………………………原頭目(Proteocophalidea)
人畜共通︰
(1)裂頭縧蟲症(Diphyllobothriasis),目前此屬縧蟲有幾個品種曾有感染人的報導,D.
latum、阿拉斯加之D.
dalliae Curtis及Bylund, 1991;北歐及加拿大之D.
dendriticum Curtis & Bylund 1991;發生於秘魯D.
pacificum Nybelin, 1931. 發生於亞洲之D.
nihonkaiense Ando et al, 2001; 北加拿大及阿拉斯加之D.
ursi Margolis et al, 1973;據1999年Crompton報導,全世界裂頭縧蟲症估計有9百萬病例。
(2)Sparganosis,曼氏迭宮縧蟲(
Spirometra mansoni)感染人類;與裂頭縧蟲區分在於第二中間宿主廣泛,不一定是魚類,尚可包含兩棲、爬蟲及某些哺乳動物。人類感染原因( i )喝入含有帶原尾呦之劍水蚤生水( ii )生吃被裂頭呦(plerocercoid, sparganum)感染之兩棲類、( iii )將被裂頭呦感染的青蛙肉、或蛇肉治療傷口。
2.病原分類︰
扁平動物門(Platyhelminthes)、縧蟲綱(Cestoda)、縧蟲亞綱(Cestoidea)、真縧上蟲目(Eucestoda)、假葉目(Pseudophyllidea)、頭槽科(Bothrioecphalidae)、頭槽縧蟲屬(Bothriocephalus);或裂頭科(Diphyllobothriidea)、裂頭縧蟲屬(Diphyllo bothrium)、舌狀縧蟲屬(Ligula)、雙線縧蟲屬(Digramma)
命名沿革︰
(1)B.
acheilognathi
B.
opsariichthydis Yamaguti, 1934; B.
fluviatilis Yamaguti, 1952; B.
gowkongensis Yeh, 1955; B.
phoxini, Molnar, 1968;
Schyzocotyle fluviatilis Akhmerov, 1960.
(2)L.
intestinalis
Fasoiola intestinalis Linnaeus, 1758; 還有其他較早文獻記載老的命名,參考Dubinina, 1966.
(3)D.
latum
Dibothriocephalus latus, Diphyllobothrium latum Linnaeus, 1758; Luhe, 1910.
疾病特性
(1) 廣節裂頭縧蟲(
Diphyllobothrium latum)之成蟲體主要寄生於犬科食肉動物,也可寄生於人,其裂頭呦(pleroercoid, sparganum)寄生於各種魚類。
一般蟲體長達10公尺,最寬處20mm,具有3,000至4,000個節片;頸節細小、呈湯匙狀、長2至3mm、寬0.7~1.0mm,蟲體背、腹側各有一條較窄而深凹之吸槽,頸部細長;成節的寬度顯著大於長度,睪丸數為750至800個,雄生殖孔和陰道外口共同開口於節片前腹面之生殖腔;子宮盤曲呈玫瑰花狀,開口於生殖腔之後,妊娠節片(gravid proglottids)之結構與成節基本相同。
蟲卵呈卵圓形,大小為58~76×40~51μm,呈淺灰褐色,卵殼較厚,一端有明顯的卵蓋,另一端有一小棘;蟲卵內含有一個卵細胞和多數卵黃細胞;排出體外時,卵內胚胎已開始發育。
蟲體之生活史,最重要的是第二中間宿主為各種魚類,人是終宿主。
一般成蟲寄生於人及犬、貓、熊、狐與豬等吃生食(生魚肉)之肉食動物小腸內;蟲卵隨宿主糞便排出後,在15~20℃水中,經過7~15天發育,孵出鈎球呦(coracidlum)。鈎球呦能在水中生存數日並能耐受一定低溫。當鈎球呦在水中生存數目並能耐受一定低溫;當鈎球幼被劍水蚤類(
Diaptomus spp.)橈腳蟲吞食後(約有40種橈腳類如Eudiaptomus及Cyclops,當為第一中間宿主),在橈腳類血腔內經過2~3週之發育為原尾呦(procerocoid);而感染縧蟲幼蟲之劍水蚤被魚吞食後,原尾呦隨血循環至魚體肌肉、性腺、肝及卵等內臟發育為裂頭呦(plerocerocoid);經宿主食入帶裂頭呦之魚時,裂頭呦在腸內經5~6週發育為成蟲;成蟲可在終宿主體內存活5~13年,見圖4。
成蟲 蟲卵 鈎球呦(coraci dium) 原尾呦(procerocoid)
(人、犬、貓等) (糞、水中) (劍水蚤吞食)
裂頭呦(plerocerocoid) 成蟲
(魚經口吞食)
(2)腸舌狀縧蟲(
Ligua intestinalis)
為甚富肌質的縧蟲,呈白色帶狀,俗稱面條蟲;長可達400mm或更長、寬7~8mm;頭節略呈三角形,身體沒有明顯的分節,其裂頭呦在背腹面中線各有一條凹陷的縱槽;成蟲寄生於食魚鳥類(如鷗類)、原尾呦寄生於鯉科魚類體腔。
蟲體之生活史為蟲卵隨糞便排入水中,孵出鈎球呦,而鈎球呦在水中游泳,被細鏢水蚤(
Diaptomus spp.)吞食後,在其體內發育為原尾呦,魚吞食帶有原尾呦之水蚤後,原尾呦穿過腸壁在魚體腔發育為裂頭呦;隨後被裂頭呦感染之魚體,再被吃食魚之鳥類吞食,如鷗(yellow herring gull,
Larus cachinnans)、(cormorant,
Phalacrocorax carbo),裂頭呦在水鳥腸中發育為成蟲,見圖3。
(3)鱊頭槽縧蟲(
Bothriocephalus acheilognathi, 同義於B.
gowkongensis Yeh, 1955)
頭槽縧蟲為一大屬,約有200餘種;蟲體呈帶狀,長20~230mm,頭節有一明顯的頂盤和2個較深的溝槽,無頸部、外分節完全。睪丸球形,每一節片約有50~90個不等,分佈於節片兩側;陰莖及陰道孔共同開口在生殖腔內,子宮彎曲呈S狀,卵楕圓形,淡褐色、殼厚蓋小,平均0.053×0.0364mm(46~48×32~34μm, Korting 1975, 50~52×33~37μm, Molnar & Murai, 1973)。
蟲體生活史經過卵、鈎球呦、原尾呦、裂頭呦及成蟲五個階段;卵在水溫28~30℃水中要3~5天始能孵化完畢、14~15℃時則需10~28天;鈎球呦圓形,後端有鈎3對,呈鐮刀狀、外膜具有纖毛,一般可在水中生活2天,在此階段內可被劍水蚤類(
Mesocyclops leuckarti)或溫劍水蚤(
Thermocyclops taihokuensis)吞食。鈎球呦穿過這些橈腳類之腸道至體腔中,約5天發育為原尾呦;原尾呦體長,尾端具有一球形尾器,內尚有3對原來小鈎,原尾呦在中間宿主體內生活時間的長短,決定於劍水蚤的壽命。帶有原尾呦之劍水蚤被魚類吞食,原尾呦就在魚腸中發育為裂頭呦;此時蟲體不分節,在夏天經11天開始長出節片,發育為成蟲;水溫28~29℃時,裂頭呦經21至23天達性成熟,開始產卵。
(4)其他縧蟲
a.鯉蠢目(Caryophyllidea)
為小型縧蟲、體不分節,僅有1套生殖器官,頭節簡單,具有淺溝淺吸盤,不具排列小鈎;附著器不發達,或具吸溝,卵有蓋、具未分裂的胚;其生活史中以淡水寡毛類(freshwater oligochaete, 顫蚓, tubificid)為中間宿主,主要寄生於淡水硬骨魚類,特別是鯉形目(歐亞及美洲)、鯰形目(亞洲魚類);約有45屬,140種縧蟲。
鯉蠢縧蟲(
Caryophyllaeus fimbriceps)為養殖於東歐及前蘇聯之鯉魚(
Cyprinus carpio)重要的寄生蟲、蟲體長15mm,具有典型頭節及排列的深溝,某些鯉蠢屬(Caryo phyllaeus)縧蟲寄生於印度及東南亞(爪畦)之鯰科魚類。
其中間宿主為環節動物的顫蚓,原尾呦在顫蚓的體腔內發育、呈圓形,體長1~5mm,前邊有1個吸附的溝槽,當魚吞食染有幼蟲之顫蚓後被感染並發育為成蟲。
另一種許氏縧蟲稱為中華許氏縧蟲(
Khavia sineusis),體長至120mm,頭節呈深切面狀形成胚胎樣頭節,發生來源於東亞,為草食性魚類如草魚(grass carp,
Ctenopharyngodon idella),此蟲分佈於許多歐洲國家及前蘇聯,同時亦寄生於中國大陸之鯉魚,分佈於湖北、黑龍江和江蘇太湖區。再者具有許多縧蟲類之紐帶縧蟲屬(Lytocestus),寄生於胡子鯰魚(
Clarias batrachus)腸,發生在印度、東南亞及非洲,此屬縧蟲的特徽於具有未分化的頭節及後卵巢區缺乏卵黃腺濾泡(vitelline follicles)。
b.窄溝目(Spathebothriidea)
蟲體長紡錘形或帶狀、背腹扁平、多節,鏈體中具線性排列的多套生殖器官,無明顯的外分節,頭節明顯成不明顯,明顯者具1或2個吸盤狀附著器官。
主要寄生於淡水及海水魚類;此類縧蟲中之截形杯頭縧蟲(
Cyathocephalus truncates),其頭節與鏈體間由一弱收縮相區別,上具漏斗狀附著器(funnel-shaped apical adhesive organ),同時俱有20~45組生殖器官,普遍寄生於鮭科魚類,與某些分佈於歐洲、北美洲及前蘇聯之淡水魚類,亦分佈於中國大陸、黑龍江流域。中間宿主為端足類(amphipods)。
c.假葉目(Pseudophyllidea)
此目縧蟲寄生於多數魚類(淡水及海水魚類),同樣亦可寄生於兩棲類、爬蟲類、鳥類及哺乳物;約有6科,60屬、280種縧蟲。也是縧蟲中的一大目;中至少有6科分類;蟲體第一中間宿主為甲殼類,第二中間宿主通常為魚類、偶見於其他脊椎動物,成蟲主要寄生於魚類,亦可寄生於其他脊椎動物︰
( i )星加縧蟲屬(Senga)
頭節呈矩形、溝槽淺但邊緣發達,頂盤背面呈鋸齒狀、鈎小排呈2個半圓形,無頸部,外分節存在,但不完全,無卵蓋、不具胚。中國大陸記錄,種鰽星加縧蟲S.
ophicephalina,寄生於烏鱧,另外在半刺光唇魚、黃鱔及鯽腸道均有被生之報導。
( ii )三枝鈎縧蟲屬(Triaenophorus)
其頭節具帶鈎的頂盤(盤上有4個三叉形的小鈎(four trident-shaped hook)、吸槽淺(具2條淺溝槽)、頸部缺、鏈體橫向皺疊、但缺外分節、具有卵蓋、含胚與否視溫度而定;主要寄生於內食性淡水魚類(特別是狗魚屬,
Esox spp.)代表蟲體為結節三枝鈎縧蟲(T.
nodulosus)及厚三枝鈎縧蟲(T.
crassus);其裂頭呦寄生在許多淡水魚類(鱸形目及鮭科魚類)之肌肉或肉臟器官(肝、腸系膜)。
d.原頭目(Proteocephalidea)
主要寄生淡水魚類,亦可寄生兩棲及爬蟲類。為小型縧蟲、頭節具4個杯狀吸盤,有時又有第五吸盤為頂吸盤、分節通常明顯、生殖孔於邊緣、卵黃腺濾泡狀、在體側或環繞節片。卵具肧,成蟲寄生於魚類、兩棲及爬蟲類,一般淡水魚被此蟲寄生之文獻記載有1科6屬;但總數有2科,約50屬,320種縧蟲。
本目代表蟲為原頭科(Proteocephalidae)內之恆河縧蟲屬(Gangesia)及原頭縧蟲屬(Proteocephalus)一般恆河縧蟲類(
Gangesia spp.)頭節上之頂突具有1~2圈鈎子,後端成熟節片長寬近等或稍長、睪丸連續、位於排泄溝之間,卵巢位於卵黃腺與節片後端間、寄生於亞洲及歐洲鯰科魚類,在中國大陸之記載有少睪恆河縧蟲(G.
oligorchis)寄生於黃顙魚,分佈於福州,鯰恆河縧蟲(G.
parasiluri)寄生於大口鯰、雜種鯰屬鯰魚,分佈於黑龍江流與遼河流域。
原頭縧蟲類(Proteocephalus)其頭節不具鈎,含有4個側吸盤、頂吸盤存在或缺;妊娠節片大於長或長大於寬、卵黃腺位於髓部排泄外側。卵呈球形、具胚;幼蟲寄生於甲殼類之血腔中發育。成蟲寄生於淡水魚類、兩棲類及爬蟲類。這屬縧蟲種類很多,已有數十種記載,分別寄生於鮭鱒魚類、鯰魚類、狗魚及江鱈等多種淡水魚類;體長6~50mm不等;大多中間宿主為劍水蚤、某些魚類受原頭縧蟲之感染程度;隨著魚體的生長發育而變動,與食性的轉變有關。
歐洲養殖之鱒魚常被P.
neglectus、短小原頭縧蟲(P.
exiguous)分佈於歐洲及亞洲,中國大陸記載寄生於鮭腸中,分佈黑龍江流域。其他長頭原頭縧蟲(P. longicollis)、小扭原頭縧蟲(P.
torulosus)亦分佈於黑龍江流域。
疾病型態及流行病學
(1)廣節裂頭縧蟲(D.
latum)
依其蟲體生活史,當哺乳動物吞食帶有裂頭呦之淡水魚類而受感染,3至6星期發育為成蟲。裂頭呦在魚類中寄生有季節性,春天多在內臟、秋天則多在肌肉,幼蟲在一般冰藏魚肉內能保持感染性達40多天,在水裡可活幾小時至幾天,在死魚肌肉內仍可活一段時間,有時一條大魚感染的裂頭呦可達1,000多個。
一般人及其他吃魚的哺乳動物(ichthyophagus mammals),吃食未煮熟(感染裂頭呦之魚類,特別是蟲體寄生於肌肉)的魚肉而感染;裂頭呦每天可長5~20cm,感染1個月後,發育為成蟲(寄生人體之蟲體,最長可至10m)。含裂頭呦之生魚肌肉可在55℃以上水溫煮5分鐘或生魚肉在零下10℃經8至72小時冷凍殺死幼蟲;但薰煙無法殺死裂頭呦;在歐洲法語區吃生魚片之食物稱為carpaccio d'mble chevalier或poissons du lac facon nordique、北義大利區稱carpaccio di persico、而亞洲地區則以日本生魚片sushi為人主要感染來源。
廣節裂頭縧蟲主要分佈在歐洲(如芬蘭、法國、義大利,前蘇聯)、美洲、亞洲(日本、大陸、台灣)之亞寒地及溫熱帶地區,俄羅斯人感染率最高,約佔全世界此病之50%以上,在人群中感染率最高的是北加拿大愛斯基摩人(83%)、前蘇聯(27%)、芬蘭(20~25%);大陸東北、廣東及台灣亦有病例報導。人體感染源都是由於誤食了生或未熟含裂頭呦之魚肉所致,喜歡吃生魚、或用少量鹽腌、薰烟之魚肉或魚卵、果汁浸魚,以及在烹製魚過程中嘗味等都易感染。流行地區人糞污染河、湖等水源使劍水蚤受污染也是一個重要因素。
本蟲體之裂頭呦感染甚多淡水魚類,包括狗魚、鱸魚;歐洲地區之斑鮭魚類(charr,
Salvelinus alpines)、江鱈魚(burbot,
Lota lota)、鱸魚(perch,
Perca fluviatilis)、狗魚類(pike,
Esox lucius)、白鮭魚類(lake,
Coregonus fera)、虹鱒(rainbow trout,
Onchorhyn- chus mykiss)、湖鱒(laek trout,
Salmo trutta lacustris)等。
(2)腸舌狀縧蟲(L.
intestinalis)
舌狀縧蟲普遍感染 屬魚類(
Barbus spp.)、鯉科魚類之鯉鯷魚(
Rastrineobola=
Engrauliccypris argenteus)、慈鯛魚類之單色鯛魚(
Haplochromis spp.)、奧利亞吳郭魚類(
Oreochromis spp.),其裂頭呦具有限制性分化結構、扁平、未分節及前端邊緣處含有2個口及溝(bothridia)、且隨寄生宿主之不同而呈現體長之差異(67~245mm×3~10mm)、且可在魚體生活達2年之久,而帶裂頭呦之魚體,被嗜魚肉之水鳥吞食(鷗,gull, cormorant),並在水鳥腸內發育為成蟲。當成蟲成熟並排而隨糞便排出體外、進入水中,卵發育呈鈎球呦,而在水中漂游,然後被水中橈腳類Acanthocyclops、Eucyceops、Diaptomus等吞食,在這些水蚤血腔發育經9~10天發育為原尾呦(視水溫而定),再經3至5天的發育,形成具感染性原尾呦;然後攜帶感染性原尾呦之淡水橈腳類被魚吃食後,則移行至宿主體腔發育,約經1年成為成熟裂頭呦,此裂頭呦可存在宿主3年甚至更久,而被裂頭呦感染的魚體,經水鳥吃食後(一般為鷗類;但亦有報告鴨、鵝、grebes、herons、egrets及ternts,亦可感染)則在腸道40~50小時內迅速發育成熟。
本蟲體分佈於全北區(Holarctic region︰包括新、舊世界北部、或古北區與新北亞區,如法國、英國、德國、波蘭、北愛爾蘭、墨西哥、捷克、加拿大、俄羅斯、突尼西亞及美國、南非、烏干達等國家)、澳亞區(Austral-asia,如中國、印度、紐西蘭、斯里蘭卡等國家);所感染的魚類,一般為以小型鯉科魚類為主要感染魚種,單就加拿大地區之報告就有30種以上魚類如下口鯉屬(Catostomus, C.
commersoni、C.
macrocheilus)、銀花鮈屬(Notropis; N.
atherinoides, N.
cornutus, N.
emiliae, N.
heterotepis, N.
hudsonius, N.
stramineus)、斑帆鮈屬(Semotilus, S.
corporalis, S.
atromaculatus)、鯉魚屬(
Cyprinus carpio)、球臉鮈屬(Hybopsis, H.
storeriana)、肥頭鮈屬(Pimephales, P.
notatus、P.
promelas)、小鰭鱸屬(Micropterus, M.
dolomieui)等,此外尚有鮭科魚類(Salmonidae)、下口鯉科(Catostomidae)、乳魚科(Galaxiidae)、棘臀鱸科(Centrarchidae)、杜父魚科(Cottidae)、棘魚科(Gasterosteidae)及河鱸科(Percidae)魚類被舌狀縧魚蟲裂頭呦寄生之報導,Loot et al. ( 2006 )建議潛在宿主之豐富性及發生強力的局部宿主專一性在於溫度的動力;而Hypsa et al. ( 2005 )提出在南非原頭目縧蟲具有前述在演化上主豐富性受溫度影響的類似證據。
(3)鱊頭槽縧蟲(B.
acheilognathi,同義於九江頭槽縧蟲B.
gowkongensis Yeh, 1955)
經由成蟲排卵,並在合適溫度下發育為纖毛幼蟲期之鈎球呦,在水中漂游狀況下,被游離自由生活之劍水蚤橈腳類(Mesocyclops、Thermocyclops、Ectocyclops、Paracyclops、Cyclops)吞食並在血腔中發育為原尾呦,然後由適量宿主吃食,然後經原尾呦在宿主腸道內發育為成蟲。此蟲體的生活史對於水溫有相當高度的依賴性;據報導在24℃下,卵發育為鈎球呦需24小時;在28~29℃下,從原尾呦發育至成蟲需21~23天,如在15~28℃水溫,則需1.5~2個月。於溫水期,完全生活史歷時1年,而於寒冷水溫蟲體發育延緩需2年或更久些;水溫在12℃下,其蟲體生活史完全被破壞停止。
本蟲體又稱為亞洲縧牋(Asian tapeworm)分佈甚廣,感染甚多魚種,包括鯉科(Cyprinidea主要感染魚種)、胎鱂亞科(Poecilidae)、慈鯛科(Cichlidae)及棘臀鱸科(Centrarchidea)、鯰科(Siluridae)、鰍科(Cobitidieae)及銀漢魚科(Atherinidae)等魚類,如 鯉魚,鲃屬(
Barbus kimberleyensis、B.
trimaculatus)、其三鼻克尼羅河魚(
Oreochromis mossambicus)、錦鯉(koi carp)、三層鯛屬(
Tritramella spp.)、食蚊鱂屬(
Gambussia affinis)、
Mirogrex terrae sanctae及
Barbus spp.。
本病原最初源自中國及日本,後迅速分散廣佈感染;原本於1954~1962發現自Amur河(Amur river)之草魚(grass carp,
Ctenopharyngodon idella),並隨即感染養殖魚群(歐洲及中國鯉魚),同時發現在亞洲與前蘇聯歐洲地區之多種野生魚類亦被寄生感染;後在1970~1975年間,分別從匈牙利、南斯拉夫、東西德報導此蟲的行蹤;至1980年後感染分佈地區更廣如法國、英國、南非、美國、墨西哥;亞洲地區如伊拉克、以色列(khalifa, 1986)、馬來西亞、斯里蘭卡(Fermando & Furtado, 1964、1963)及韓國(kim et al., 1985)陸續發現。相同屬不同種之蟲體如發現於埃及之B.
aegypticus (Amin, 1978)、發現於中南非之B.
kivrensis(Baer & Fain, 1960; Mashego, 1982)。
亞洲縧蟲似乎是一種溫度專一性品種(thermophilic species),低溫下似乎可延遲或阻礙蟲體的生長,在28~30℃水溫下,5天內約有77%的蟲卵可孵育;而水溫14~15℃,其蟲卵之孵育時間延長至10~28天。而鈎球呦可被廣泛淡水橈腳類吞食,亞洲地區以
Mesocyclops leuckarti最普遍,在中國及韓國則記載有
Thermocyclops spp.、
Ectocyclops spp.及
Paracyclopos spp. 三種橈腳類吞食本蟲卵。在28~29℃水溫下,本蟲體之原尾呦發育至成蟲需21~23天,而在15~22℃水溫時,則需1.5~2月;水溫15℃以下,則更延長至6~8月;在寒冷水溫下完整的生活史;需經2年或更久的時間。
亞洲縧蟲成蟲之蟲長似乎與寄生宿主體長大小有關,寄生於幼鯉魚(體長100~150mm)及草魚之成熟蟲體有500~600節片;T.
canis體長大於60mm及慈鯛魚類體長大於40mm,成蟲亞洲縧蟲至少有100個節片及150~350之妊娠節片,最長至580個節片。而體長小於50mm之T.
canis及體長小於35mm之慈鯛魚類,成蟲體節片少於60個節片及妊娠節片少於100個節片;再體長更短(17~20mm)慈鯛魚類之魚苗,成熟蟲體之節片及妊娠節片合計少於50個節片,甚至可少於17~20個節片。
亞洲縧蟲感染地區分佈很廣,包括歐洲、亞洲、非洲及美洲;歐洲地區有法國、義大利、匈牙利、捷克、德國、英國、奧國、南斯拉夫、前蘇聯;亞洲地區有中國、日本、韓國、菲律賓、斯里蘭卡、馬來西亞、以色列及伊拉克等,美洲如美國、墨西哥、加拿大;非洲則以南非為主要地區。
(4)其他縧蟲
各地區養殖及野生魚類感染的縧蟲各有其品種上的差異性,以非洲地區不同種類成熟縧蟲至少超過40種寄生魚類之縧蟲。一般以鯉蠢目(Caryophyllaeidae)(無分節、屬monzoic)如同假葉目及原頭目(有分節、屬polyzoic)樣俱雙棲特性;鯰形目(Siluriform)魚類通常是無分節及有分節縧蟲的共同宿主。
養殖及野生海水魚類之海水真縧蟲(marine eucestoda)有三個族羣,假葉目、四葉目(Tetraphyllidea)及鍾吻目(Trypanorhynchidea = Tetrarhynchidea),而主要區別特徵在於頭節構造,而尚有少許品種屬於雙葉目(Diphyllidea)、原頭目、盤頭目(Lecanicephalidea)及窄溝目(Spathebothriidea)︰
a.錐吻目(Trypnorhynchidea)︰主要寄生於板鰓魚類、第一中間宿主為甲殼橈腳類,可能為海水非脊椎動物、第二中間宿主為海水硬骨魚類,具有19科、50屬,約300~350種縧蟲。
一般頭節可分四區,突盤部、鞘部、球體部和球體後區,具2或4個通常無柄的突盤和4個可外翻的錐吻,吻上具形態、數目、排列各異的鈎;吻鞘後部為一長形肌肉球。幼蟲寄生於魚類、海產無脊動物,少數寄生於爬蟲類;成蟲寄生於板鰓魚類(elasmobranch fishes)的胃或螺旋瓣中。
一般原尾呦寄生在浮游性橈腳類,裂頭呦寄生在海水魚類及十足目(decapods)體腔或肌肉、成蟲寄生於板鰓類魚類腸道,而
Gilquinia squali之裂頭呦則寄生於宿主眼睛;如同墨西哥灣裏之
Prochristianella hispida以蝦為中間宿主,典型常見包括Dasyrhynchus、古力羅縧蟲(Grillotia)、尼柏縧蟲(Nybelinia)、Gymnorhynchus、Hepatoxylon、Lecistorhynchus。
b.四葉目(Tetraphyllidea)—主要寄生於板鰓及全頭魚類(holocephalan fishes),第一中間宿主未明、幼蟲寄生於海水硬骨魚類(可能為第二中間宿主或伴侶性宿主),具7科、60屬,約800種縧蟲。
一般蟲體中等大小、頭節有4個葉狀、喇叭狀或耳狀之突盤(裂片)(four prominent leaf-like outgrowths),分節通常明顯,為覆疊鏈體或非覆疊鏈體(acraspedote),妊娠子宮粗大、占據節片大部份;卵內具六鈎呦;寄生於海水魚類、兩棲及爬蟲類消化道。生活史大部份尚未清楚,原尾呦相信寄生於浮游性橈腳類,裂頭呦寄生魚體腸道,發育為成蟲後,寄生於板鰓類魚體腸道;此目中某些種類縧蟲之裂頭呦特別被稱為Scolex pleuronectis及Scolex polymorphus;典型種類有鈎槽縧蟲屬(Acanthobothrium van Beneden, 1849)、葉槽縧蟲屬(Phyllobothrium Beneden, 1849)及鮣槽縧蟲屬(Echeneibothrium Beneden, 1850),個別蟲體很少超過10cm,具有3種不同方式附著吸附於板鰓類魚之腸道中。
c.假葉目(Pseudophyllidea)—主要寄生於淡水及海水硬體魚類,同時亦可寄生於兩棲、爬蟲、鳥類及哺乳動物;中間宿主為甲殼類、第二中間宿主為魚,很少為其他脊椎動物,具6科、60屬,約280種縧蟲。
頭節過常存在,上具背腹兩吸槽,鈎有或無;頭節偶有付缺或為假頭節或特化頭節(Scolex detormatus)所替代;吸槽偶爾付缺,外分節一般存在,完全或不完全,偶爾付缺。第一中間宿主為甲殼類、第二中間宿主通常為魚類,成蟲主要寄生於魚類,亦可寄生於其他脊椎動物。
溯游性魚類,主要為鮭科魚類,常從海中或淡水區域攜帶大量縧蟲,裂頭呦寄生於魚體肌肉(如改變外在環境鹽度,則影響裂頭呦寄生),多數裂頭縧蟲(
Diphyllobothrium spp.)成熟寄生於人體;另外同屬不同種之D.
pacificum亦可發現寄生於太平洋海獅或海豹(fur seal),另外
Diplogonoporus balaenoptera除了寄生於多數品種鯨魚腸道外,日本Kamo et al. ( 1971 )報告成蟲長約10.6m寄生於人體。典型蟲體有頭槽縧蟲屬(Bothriocephalus Rudolphi, 1808)、裂頭縧蟲屬(Diphyllobothrium Linnaeus, 1758)、Diplogonoporus屬、真溝縧蟲屬(Eubothium Nybelin, 1922)、Abothrium屬。
臨床症狀及病理學
(1)廣節裂頭縧蟲(D.
latum)
因為裂頭呦感染魚體之肌肉,一般在臨床上並無明顯之特異性病變,蟲體可在肌肉形成囊疱,所以必需檢查被寄生之魚體肌肉是否呈現白色囊狀病變。如果感染人體,因蟲體寄生腸道,多數感染者無明顯臨床症狀;少數人呈現疲勞、乏力、四肢麻木、腹瀉或便秘以及飢餓感,嗜食鹽等較輕微症狀。但因蟲體長大,有時蟲體可扭結成團,導致腸道、膽道阻塞,甚至出現腸穿孔等症狀。此外,尚有人肺部和腹膜發現廣節裂頭呦寄生之報告;約有2%的廣節裂頭縧蟲病人併發縧蟲性貧血,可能與造血功能有關的Vit B12被縧蟲大量吸收,如果食物中Vit B12供給不足,則可能引起Vit B12缺乏;另外縧蟲的代謝產物可能損害宿主造血功能;患者除有一般惡性貧血的表現外,常出現感覺異常、運動失調、深部感覺缺失等神經紊亂現象,嚴重者甚至失去工作能力,驅蟲後貧血能快好轉。
(2)腸舌狀縧蟲(L.
intestinalis)
因為蟲體原尾呦穿過腸壁到達體腔並發育為裂頭呦,造成病魚腹部膨大,嚴重時失去平衡,側游上浮或腹部朝上;剖檢時,可見病魚體腔中充滿大量白色帶狀之蟲體;內臟因受蟲體擠及纏繞,形成諸臟器如肝、脾、性腺及腸變形萎縮,臟器正常功能受損,引發生長遲緩、魚體消瘦、嚴重貧血、無法生殖。有時裂頭呦尚可從魚腹部鑽出,直接造成幼魚死亡。病理變化,可見組織纖維化、炎症反應、血液樣腹水蓄積、貧血,由於寄生蟲壓擠形成內臟器官萎縮;因為長期飢餓,導致宿主缺鐵、葉酸和Vit B12等物質,同時影響白血球正常組成變化,據Mazur & Pronin ( 2006 )報導此蟲體感染泥鰍(
Rutilus rutilus lacustris)會降低紅血球及白血球總量、增加泡沫細胞(foam cells)、抑制T淋巴球相關免疫反應、降低總蛋白、白蛋白及r球蛋白分片之含量;據研究指出,由於體腔中的裂頭呦能釋出某種物質,使機體產生免疫反應,但不同宿主,寄生期以及不同的寄生蟲可能產生不同的差異反應。
(3)鱊頭槽縧蟲(B.
acheilognathi)
臨床可見病魚腹部膨脹、腹水蓄積、體重減輕、體色變黑、離群獨游,並伴隨貧血,被感染的魚種紅血球降低;嚴重感染時,前腸第一盤曲脹大成胃囊狀,直徑增加3倍;腸壁皺褶萎縮,表現出慢性炎症反應;組織病理的表現與寄生宿主品種、大小及條件有所差異,包括寄生蟲體之壓力擠壓臟器、腸道發炎,尤是蟲體附著處呈現嚴重卡它出血性腸炎(catarrhal hemorrhagoc enteritis),並促使周圍結締組織增生;Hoffman(1980)報告golden shiner之小魚被本病原寄生時,腸道可能阻塞甚至形成腸穿孔之病灶。魚體之死亡率與蟲體寄生強度及魚體大小有關(實驗證明亦有相同結論);Bauer et al.(1969)報導嚴感染幼鯉魚,其死亡率可達90%;而Scott & Grizzle(1979)發現被本病原感染的草魚及golden shiner(
Notenigonus crysoleucas)很少提起病理變化,因為病灶局限在被蟲體吸附之處;同時發現感染蟲體與未感染蟲體之魚體血容比值(haematocrit)類似,蟲體感染的病理效應正確的評估相關不但是於蟲體破壞宿主腸壁組織,而且決定於宿主大小與各種條件的伴隨的因素;雖然亞洲縧對魚種並無嚴格的宿主專一性寄生,但確有對不同宿主品種之差異性的感受性寄生,這與被感染魚體之防禦反應有著不同的變異反應;Scott et al.(1979)證實亞洲縧蟲的寄生具有機械損傷作用;腸粘膜下層淋巴細胞增多,有些於寄生處呈現出血現象, 草魚的腸粘膜淋巴細胞浸潤不在溝槽吸著處,而出現於其他部位病灶處。
在中國大陸亞洲縧蟲可造成養殖草魚大量死亡,每年育苗初期間開始感染,在短期內,大都魚種病情趨於嚴重,越冬草魚之死亡率可達90%;但魚種體長超過10cm時,感染率下降;2年齡以上之魚群,僅偶有少數頭節和不成熟的幼體,這與草魚在不同發育階段食性的改變有關。
(4)其他縧蟲
寄生海水魚類之縧蟲大部份歸屬於錐吻目、四葉目及假葉目︰
a.假葉目—
Bothriocephalus scorpili
感染東北大西洋海域內之sand goby,其裂頭呦寄生於魚體胃壁,或嚴重寄生turtot,蟲體長至60cm;大量感染導致魚體腹部膨大、刺激產卵,但無嚴重大量死亡之疫情報導。
b.四葉目—本目蟲體主要寄生於鯊魚及鰩(rays)從鈎槽縧蟲屬(Acanthobothrium van Beneden, 1849)為代表,一般頭節呈錐形或鱗莖狀,突盤延長、卵形或圓形,各自被二橫隔分成3個小室,前端具2枚雙叉形鈎,鈎的正前方為一小附加墊或附加吸盤,具頸部。前部的節片排列緊密,後部節片寬大於長或長大於寬,可能略帶覆瓦狀、或節片間略有收縮,卵小,呈亞球形。有廈門鈎槽縧蟲(A.
xiamenensis)寄生於達尖梨頭鰩,分佈於福建海區;冠鈎槽縧蟲(A.
coronatum)寄生於赤魟,分佈福建平潭,大頭鈎槽縧蟲(A.
grandiceps)寄生於尖嘴魟、赤魟,分佈於東海。
c.錐吻目
引起大量經濟損失,是由於不同錐吻目縧蟲之裂頭呦大量寄生魚體肌肉,幼蟲寄生於脊椎附近;某些種類個體之長度可達20cm。以常見的古力羅縧蟲屬(Grillotia)為例,裂頭呦寄生於真骨魚類的腹膜或肌肉中形成包囊(blastocyst),成蟲寄生於板鰓類。
一般頭節長、突盤帽貝形、後部可能具齒,吻異鈎型,具斜行橫向半回轉排列,具形狀、大小各異的鈎,鞘部長於突盤部、球體部很長,牽引肌插入球體的內腔。蟲卵具卵圓或橢圓球狀,可能有一短極絲。
寄生於板鰓類的有
Grillotia meteori sp. nov., G. cf.
microthrix,未曾有引起大量經濟損失之報導。
病原致病性意義
腸舌狀縧蟲(L.
intestinalis)寄生引發的疾病對於商業性養殖之鯉科魚類及捕獲鯛魚(
Abramis brama)帶原是一項嚴重的問題,由於某些帶原魚類被腸舌狀縧蟲寄生,引發魚產量的下降,並減少魚體的商品價值。尤其是中國大陸地區隨著淡水養殖業的飛速發展,特別是大水面淡水養殖病情更加嚴重,因為此蟲體之裂頭呦可寄生多種魚類(鯽、鯉、鰱、鱅、鯿及鮊等魚種),對寄主無宿主專一性寄生,對於1年齡鯉科魚類的感染相當嚴重;因此,防治的措施需詳加研究及推演。
亞洲縧蟲即鱊頭槽縧蟲(B.
acheilognathi)感染中國大陸、前蘇聯及菲律賓養殖鯉魚非常嚴重,同時也昃美國南部魚餌工業污染此病原的嚴重問題;大部份感染於溫暖及次熱帶區域魚類。
寄生蟲對魚類的危害及影響,應是機械性刺激所造成組織的傷害,直接引發其他病變,甚至影響臟器功能,再者因蟲體的擠壓及阻塞,引發臟器萎縮、壞死及生理機能喪失;並且掠奪宿主營養,使寄生宿主營養不良、生長遲緩、無法生育;有些寄生蟲甚至會產生毒素,因寄生蟲代謝產物排泄於宿主體內,在某些情況下,對寄生機體產生一定的影響;這些均是寄生蟲對宿主的傷害之病原意義。

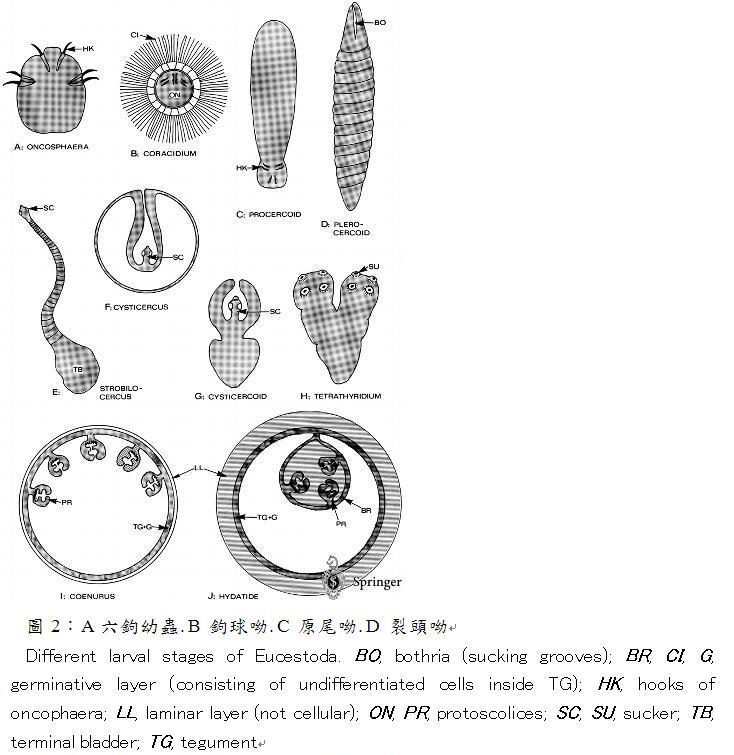
圖1:各種縧蟲頭節之構造
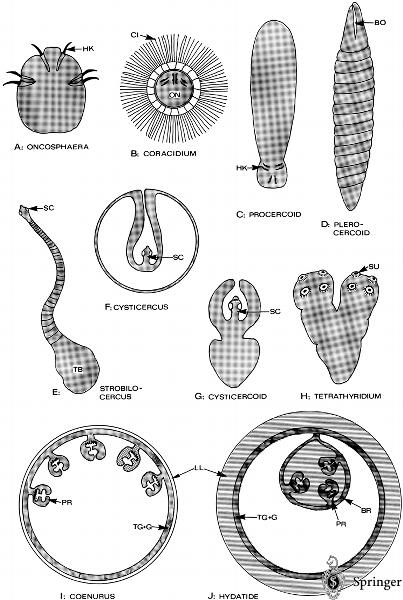
圖2:A 六鉤幼蟲. B 鉤球呦. C 原尾呦. D 裂頭呦
Different larval stages of Eucestoda. BO, bothria (sucking grooves); BR, CI, G, germinative layer (consisting of undifferentiated cells inside TG); HK, hooks of oncophaera; LL, laminar layer (not cellular); ON, PR, protoscolices; SC, SU, sucker; TB, terminal bladder;

圖3 腸舌狀縧蟲生活史
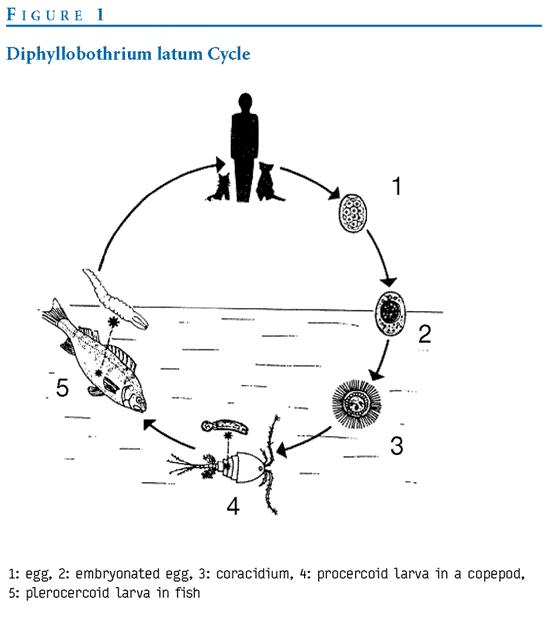
圖4. 廣節裂頭縧蟲生活史
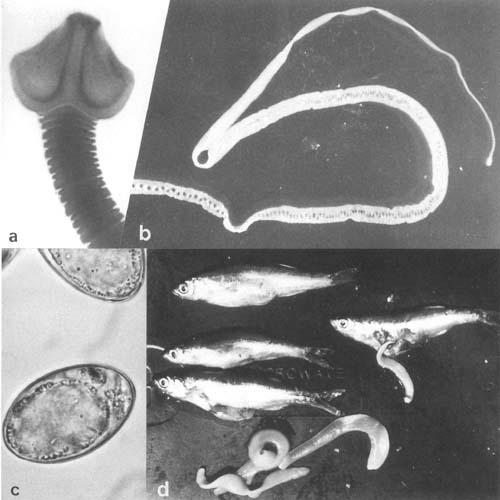
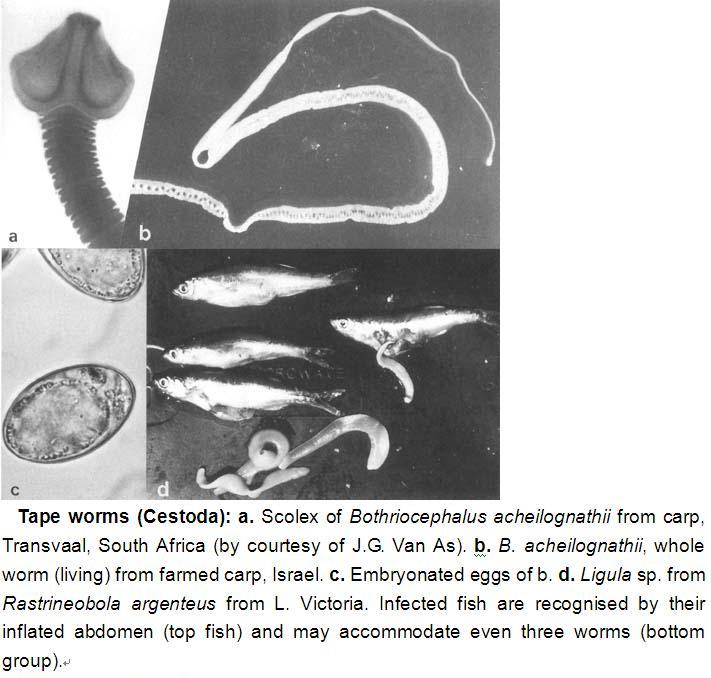
Tape worms (Cestoda): a. Scolex of Bothriocephalus acheilognathii from carp, Transvaal, South Africa (by courtesy of J.G. Van As). b. B. acheilognathii, whole worm (living) from farmed carp, Israel. c. Embryonated eggs of b. d. Ligula sp. from Rastrineobola argenteus from L. Victoria. Infected fish

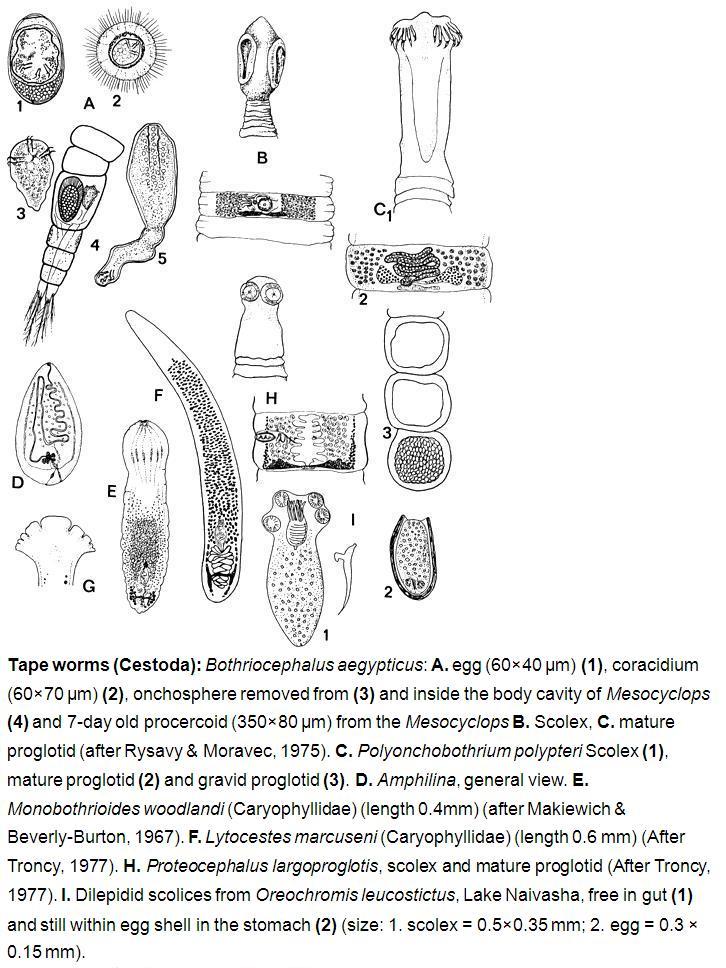
Tape worms (Cestoda): Bothriocephalus aegypticus: A. egg (60×40 μm) (1), coracidium (60×70 μm) (2), onchosphere removed from (3) and inside the body cavity of Mesocyclops (4) and 7-day old procercoid (350×80 μm) from the Mesocyclops B. Scolex, C. mature proglotid (after Rysavy & Moravec, 1975). C. P
圖片參考來源:http:// parasitology. Informatik.uni-wuerzburg.de
圖片參考來源:http: // www.fao.org/docrep/
圖片參考來源:http://www.fao.org/docrep
2. 診斷方式
一級初步診斷:
依據魚體表現的臨床症狀及肉眼病變,而自瀕死魚體所採的標本將予適當保存,便於進一步鑑定蟲名種類。
同時由鯉科魚類膨大腹部,剖檢後自體腔所得白色帶狀蟲體,呈現大型、平滑、彎曲,無分節之裂頭呦(Plerocercoid)背和腹部均具有明顯之溝槽構造,判定可能為舌狀縧蟲(
Ligula spp.)需與兩線縧蟲區分。
二級初步診斷:
在淡水魚(特別是鯉科魚類)腸道中呈現明顯心臟形(heart-shaped)頭節,推斷可能為頭槽縧蟲(
Bothriocephalus spp.);在蟲體鑑別診斷中,位於全北區(Holarctic)頭槽縧蟲品種均具有長的頭節並呈心臟形,包括B.
kivuensis(具有50~75個睪丸、卵之大小為50~54×34~36μm)、B.
aegypticus(具有140~200個睪丸,卵之大小為66×34~46μm)分佈於北非鯉科魚類。
廣節裂頭縧蟲(D.
latum)因裂頭呦寄生於魚體肌肉或體腔,且頭節具有明顯的持徵;因此,初步可以區別。
三級初步診斷:目前尚無可應用的方法
一級確定診斷:目前尚無可應用的方法
二級確定診斷:
1.依據Pool & Chubb, 1985及Paperna, 1996發表之文獻,由蟲體形態特徵及表量各組成的衡量大小,同時需與B.
kivuensis(中南非,Baer & Fain, 1958)、B.
aegypticrs(埃及,Rysavy & Moravec, 1975)區別,因為這二蟲體之頭節亦呈心臟形,也感染鯉科魚類(
Barbus spp.)。Hoffman(1999)認為前述二蟲體應與B.
acheilognathi同種異名,但Paperna(1996)認為前述二蟲體各組成之特徵可以區別;因為B.
aegypticus之睪丸具有140~200個(140~280, Aim, 1978)、蟲卵為60×34~46μm(Rysavy & Moravec, 1973),而B.
kivuensis睪丸有50~75個、蟲卵大小為50~54×34~36μm(Baer & Fain, 1958. 1960),體長700mm至1m,為B.
acheilognathi體長的3倍,同時具有陰道括約肌(vaginal sphincter)。
2.舌狀縧蟲類(
Ligula spp.)之裂頭呦需與非常相似的兩線縧蟲類(Digramma),因為兩線縧蟲裂頭呦具有生殖複合器(genital complexes)並擴延呈單一緃列,而舌狀縧蟲相反且有2列。從裂頭呦的形態上仍是很難區別,由對宿主專一性與地理上分佈的意義也仍不明,但最近由實驗感染吃魚鳥類使蟲體發育至成熟縧蟲之觀察,是新的分類依據。
3.廣節裂頭縧蟲(D.
latum),因為人類吃食被裂頭呦感染的魚體之生魚片而感染;但這屬內不同國家地區,呈現同屬不同種的蟲體,亦同樣感染人類,所以需在形態上、表量方面仔細的度量區別。
4.其他種類縧蟲
可依據文獻記載、專門的寄生蟲學術期刊或書籍,借由蟲體頭節附著器的特徵、感染魚種的部位與分佈的地理區域或藉及掃描式電子顯微鏡所呈現的超顯微結構,特別是頭節的組成,應可明確的察明及確定品種。
三級確定診斷:應用分子生物學診斷技術
1.裂頭縧蟲(
Diphyllobothrium spp.)具有廣泛的宿主域,包括感染人類;依據Muller(2002)及Chai et al.(2005)之報導,全球每年約有2千萬的感染病例,分佈的地區甚廣包括北美北部、歐洲、俄羅斯、siberia;因為同屬不同種的裂頭縧蟲相繼被發現命名;為了更詳細區別,近年來在種方面的鑑定,採用18S rRNA內的部分核苷酸ITS2序列及細胞色素C氧化酶次單位I基因(Cytochuome C oxidase Subunit I. gene; COI)來分析彼此間差異,因為應用種專一性序列(species-specific sequence),可作為有利的例行診斷工具,不但可明確區明品種,又可迅速判定,解決處理公共衛生的問題。
2.亞洲縧蟲(B.
acheilognathi)
因為頭槽縧蟲為縧蟲內的一個大屬,約有200多品種,且寄生的宿主廣域,雖然主要為鯉科魚類,但感染縧蟲品種繁多,如依據傳統形態方法行品種鑑定將緩不待急,今拜分子生物學診斷技術的發達,在此蟲鑑定上近年來採用differential display(DD), RNA arbitrarily primer PCR(RAP-PCR)、Suppression substractive bybridization(SSH)等方法;可快速區別裂頭縧蟲的品種。
3.舌狀縧蟲(Ligula spp.)有廣泛的宿主域,可寄生於多科魚類,包括鯉魚科、下口鯉科、鮭科魚類、杜父魚科、乳魚科、棘魚科等,而終宿主為吃魚鳥類,其水鳥種類種更為繁多,均可能攜帶舌狀縧蟲之成蟲,並隨棲息地區的不同感染不同區域的魚類,而顯現的腸舌狀縧蟲有著種別上的差異,應用傳統形態學方法,無法解決舌狀縧蟲在地理分佈區及不同宿主魚類所帶來的形態差異;近年來採用全部ITS區域及28s rRNA部份核苷酸序列分析,或是使用inter-simple sequence repeat(ISSR)當為標誌來區別多數形態學上無法辨別舌狀縧蟲之裂頭呦在基因相關演變的差異分析。
3. 治療方法
目前尚無適合的治療方法,再者行政院農業委員會動植物防疫檢疫局所訂定的「水產動物用藥使用規範」裏並無列舉驅縧蟲藥,所以業者需從預防方面著手,所謂預防重於治療的原則,即來表現此時的應用狀態。
4. 預防措施
1.明瞭縧蟲的生活史,採取截段生活史的生物防治方法。
2.池塘清理時,除日常的翻晒外,在加上生石灰的處理,用來殺滅蟲卵。
3.清塘時採用20ppm漂白粉,來殺滅劍水蚤及蟲卵。
4.有條件性的儘量趨趕鷗鳥,以杜絕傳染病。
5.育苗期間為了避免污染蟲源,最好自己培養橈腳類,儘量不要到其他不明區域撈取或購買橈腳類餵食。
5. 參考資料
1.張劍英.邱兆祉.丁雪娟等編著,魚類寄生蟲與寄生蟲病.第十一章寄生於魚類的縧蟲與縧蟲病,1999,科學出版社。
2.Moller H, Andevs K. Diseases and Parasities of Marine Fishes. 4-parasites, 4.6.3-cestoda、1986, Scanner-Studio-Nord, D-2300. Kiel 14 Printed in Germany.
3.FAO corporate Document Repository.
Parasites. Infections and Diseases of fishes in Africa: An Update. 14 Tapeworm ( Cestoda ).
4.Bouzid W et al. Genetic diversity of
Ligula inetstinalis ( Cestoda: Diphyllobothriidea ) based on analysis of inter-simple sequence repeat markers. J. Zool Syst. Evol. Res. 2008; 46(4):289-296.
5.Logan FJ, et al. The phylogeny of diphyllobothrid tapeworms ( Cestoda: Pseudo- phyllidea ) based on ITS-2 rDNA sequences. Parasitology Research. 2004.
6.Luo HY, et al. Characterization of development-related genes for the cestode
Bothriocephalus acheilognathi. Parasitology Research. 2004.
7.Salgado-Maldonado G., et al. The Asia fish tapeworm
Bothriocephalus acheilognathi: a potential threat to native freehwater fish species in Mexico. Biological Invasions. 2003; 5:261-268.
8.Mazur OE, Pronin NM. Parameters of the blood and immune system of
Rutilus rutilus lacustrisCypriniformes: Cyprinidae ) during an invasion with Pleurocercoids of
Ligula intestinalis ( Pseudophyllidea: Ligulidae ). Journal of Ichthyology. 2006; 46(5):399-403.